
CIN - PowerPoint PPT Presentation
Title: CIN
1
CINÉTICA ENZIMÁTICA
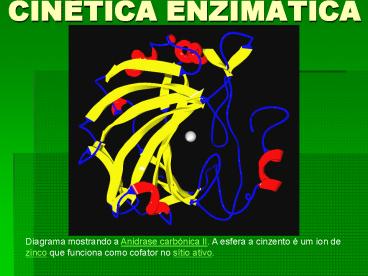
Diagrama mostrando a Anidrase carbónica II. A
esfera a cinzento é um ion de zinco que funciona
como cofator no sítio ativo.
2
CINÉTICA ENZIMÁTICA
3
CINÉTICA ENZIMÁTICA
4
A linha em vermelho representa a reação na
ausência de catalisadorA linha em azul na
presença de enzima. S nível de energia do(s)
substrato(s) P nível de energia do(s)
produto(s) G energia de Gibbs R coordenada
de reacção (sentidos de progressão de
reacção)?G'º energia livre padrão bioquímica
?G energia de ativação S-P do(s)
substrato(s) P-S do(s) produto(s)).
CINÉTICA ENZIMÁTICA
5
A cinética enzimática é o estudo do mecanismo
pelo qual as enzimas ligam substratos e os
transformam em produtos. São utilizados dados
sobre a velocidade de reação de enzimas através
de ensaios enzimáticos.
Mecanismo de reação de uma enzima com substrato
único. A enzima (E) liga o substrato (S) e
liberta o produto (P).
6
Cinética de MichaelisMenten Curva de saturação
para uma enzima mostrando a relação entre a
concentração do substrato e a velocidade de
reação As reações catalisadas por enzimas são
saturáveis, e a sua velocidade de catálise não
indica uma resposta linear face ao aumento de
substrato. Se a velocidade inicial da reação é
medida sobre uma escala de concentrações de
substrato S) a velocidade de reação (v),
aumenta com o acréscimo de S. A medida que S
aumenta, a enzima satura-se e a velocidade atinge
o valor máximo Vmax.
7
CINÉTICA ENZIMÁTICA
A saturação acontece porque, à medida que é
aumentada a concentração de substrato, aumenta
também a quantidade de enzima presente sob a
forma de complexo enzima-substrato (ES). À
velocidade máxima, Vmax, todos os centros ativos
estão ocupados (saturados) com substrato, ou
seja, não existe enzima livre para ligar mais
substrato e a concentração de complexo ES é igual
à concentração de enzima.
8
CINÉTICA ENZIMÁTICA
A atividade de uma enzima é geralmente descrita
através de Vmax e também da constante de
Michaelis-Menten (KM), que representa a
concentração de substrato à qual se detecta uma
velocidade de reacção igual a metade de Vmax
Cada enzima possui um KM característico (kcat)
para um dado substrato este parâmetro é muitas
vezes usado para demonstrar a força de ligação de
um substrato à enzima. É também usada outra
constante cinética, kcat, para descrever o
comportamento de uma enzima, representando o
número de moléculas de substrato que podem ser
catalisadas por centro ativo por segundo.
9
No modelo da cinética de Michaelis-Menten de uma
reação com um substrato existe uma reação
bimolecular inicial entre a enzima E e o
substrato S para formar o complexo ES. Embora o
mecanismo enzimático para a reação unimolecular
possa ser bastante
complexo, há tipicamente um passo na determinação
da velocidade que permite que se modele o
mecanismo como um passo catalítico simples de
velocidade constante k2.
(Eq. 1). k2 também é o valor máximo
de reações enzimáticas catalisadas por segundo.
10
Com baixas concentrações de substrato S, a
enzima permanece em equilíbrio entre a forma
livre E e o complexo enzima-substrato ES
aumentando S aumenta ES à custa de E,
mudando o equilíbrio para o lado direito.
Uma vez que a velocidade de reação depende da
concentração ES, a velocidade é sensível a
pequenas alterações de S Altos valores de S,
a enzima fica totalmente saturada com substrato,
e existe apenas na forma ES. Nestas condições, a
velocidade (vk2EtotVmax) é insensível a
pequenas alterações de S assim, Etot é a
concentração total enzimática
que é aproximadamente igual à concentração ES
em condições de saturação.
11
A equação de MichaelisMenten7 descreve como a
velocidade de reação v depende da posição do
equilíbrio ligado ao substrato e da constante de
velocidade k2. Michaelis e Menten mostraram que
se k2 for bem menor que k-1 (chamada a
aproximação de equilíbrio), pode obter-se a
seguinte equação
A constante de Michaelis Km é definida como a
concentração para a qual a velocidade da reacção
enzimática é metade de Vmax. Isto pode
verificar-se ao substituir Km S na equação de
Michaelis-Menten. Se o passo para determinação
da velocidade for lento, quando comparado com a
dissociação do substrato (k2 ltlt k-1), então a
constante de Michaelis Km é aproximadamente à
constante de dissociação do complexo ES, embora
tal situação seja relativamente rara.
12
A situação mais normal dá-se quando k2 gt k-1 na
por vezes chamada cinética de Briggs-Haldane. A
equação ainda se verifica em condições mais
gerais, como provém da aproximação ao estado
estacionário. Durante o período inicial, a
velocidade de reação v é relativamente constante,
indicando que ES é também aproximadamente
constante (cf. equação 1)
A concentração de ES é dada por
em que a constante de Michaelis Km é definida
por
13
(E é a concentração de enzima livre) . Em
conjunto, a fórmula geral para a velocidade de
reação v vem pela equação de Michaelis-Menten
A constante de especificidade kcat / Km é uma
medida da eficiência de conversão pela enzima do
substrato em produto. Usando a definição da
constante de Michaelis Km, a equação escreve-se
na forma
14
O gráfico de velocidade face a S não é linear.
Embora a baixas concentrações de substrato se
mantenha linear, vai curvando à medida que
aumenta a concentração de substrato. Antes da
chegada dos computadores, que permitem ajustar
regressões não lineares de forma simples, podia
chegar a ser realmente difícil estimar os valores
de Km e Vmax nos gráficos não lineares. Então
vários pesquisadores concentrassem os seus
esforços em desenvolver métodos de linearização
da equação de Michaelis-Menten, dando como
resultado o gráfico de Lineweaver-Burke
15
O gráfico de Lineweaver-Burk ou representação de
duplo recíproco é a forma mais comum, e
simples,de mostrar os dados da cinética
enzimática. Para tal, tomam-se os valores
inversos de ambos os lados da equação de
Michaelis-Menten. Como se pode apreciar na figura
da direita, o resultado deste tipo de
representação é uma linha reta cuja equação é e
mx c, sendo o ponto de intercecção entre a reta
e o eixo das ordenadas equivalente a 1/Vmax, e o
ponto de intercepção entre a recta e o eixo de
abcissas equivalente a -1/Km.
O gráfico de Lineweaver-Burke ou do duplo
recíproco mostra o significado dos pontos de
intercecção entre a reta e os eixos de ordenadas,
e do gradiente da velocidade.
16
INIBIÇÃO ENZIMÁTICA
Esquema cinético para inibidores enzimáticos
reversíveis. EEnzima SSubstrato ESComplexo P
Produto IInibidor
17
INIBIÇÃO ENZIMÁTICA
Inibidores enzimáticos são moléculas que reduzem
ou eliminam a atividade das enzimas. Podem
encontrar-se dois tipos Reversíveis quando a
remoção do inibidor restaura a atividade
enzimática classificados como competitivos,
acompetitivos, não-competitivos e mistos, segundo
o efeito que produzam nas constantes cinéticas Km
e Vmax. Este efeito dependerá da união do
inibidor à enzima E, ao complexo
enzima-substrato ES ou a ambos. Irreversíveis
quando o inibitor inativa de modo permanente a
enzima. Geralmente produzem modificações
covalentes de resíduos do centro catalítico da
enzima. Estas reacções decaem de forma
exponencial e são habitualmente saturáveis.
Abaixo dos níveis de saturação, mantêm uma
cinética de primeira ordem em relação ao
inibidor.
Recommended
CrystalGraphics Presentations




























![❤[READ]❤️ L'Histoire du cinéma illustrée pour les Nuls, nelle édition (French Edition) PowerPoint PPT Presentation](https://s3.amazonaws.com/images.powershow.com/10085050.th0.jpg?_=20240724059)
