
Prezentace aplikace PowerPoint - PowerPoint PPT Presentation
1 / 39
Title:
Prezentace aplikace PowerPoint
Description:
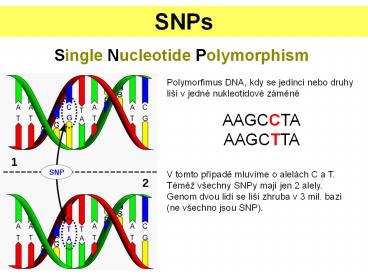
SNPs Single Nucleotide Polymorphism Polymorfimus DNA, kdy se jedinci nebo druhy li v jedn nukleotidov z m n AAGCCTA AAGCTTA V tomto p pad mluv me o ... – PowerPoint PPT presentation
Number of Views:102
Avg rating:3.0/5.0
Title: Prezentace aplikace PowerPoint
1
SNPs
Single Nucleotide Polymorphism
Polymorfimus DNA, kdy se jedinci nebo druhy liší
v jedné nukleotidové zámene AAGCCTA AAGCTTA V
tomto prípade mluvíme o alelách C a T. Témež
všechny SNPy mají jen 2 alely. Genom dvou lidí se
liší zhruba v 3 mil. bazí (ne všechno jsou SNP).
2
SNPs
3
SNPs
4
SNPs genotypizace
- Hybridizacní metody
- Enzymatické metody
- Metody založené na fyzikálních vlastnostech DNA
Hybridizacní metody
Molecular beacon
5
SNPs hybridizacní metody
906 600 lidských SNPs
6
SNPs enzymatické metody
Primer extension napr. Infinium (Illumina)
http//www.illumina.com/technology/infinium_hd_ass
ay.ilmn
7
SNP OSTATNÍ METODY
Single-Strand Conformation Polymorphism
denaturace
PCR
renaturace
vertikální elektroforéza
vizualizace
8
Srovnání metod
Metoda DNA hybri dizace Mass fingerptinting Mikro satelity RFLF (VNTR) SINE RAPD AFLP SNP
Pocet lokusu Všechny Mnoho Jeden Mnoho Jeden Mnoho Mnoho Jeden / mnoho
Repliko vatelnost Ruzná Vysoká Vysoká Vysoká Vysoká Ruzná vysoká vysoká
Povaha znaku Distance Kodominantní Kodomi nantní Kodomi nantní? Vzácná událost Domi nantní Kodominantní? Kodo minantní
Rozlišení Strední vysoké Vysoké Vysoké Nízké Strední Vysoké Vysoké
Jedno duchost provedení Težké Jedno duché Težké Jedno duché Težké Jedno duché Strední -
Doba trvání Strední Krátká Dlouhá Krátká Dlouhá Krátká Krátká -
9
rEKAPITULACE
Jak získáváme znaky pomocí sekvenace unikátních
lokusu
Sekvenace lokusu u taxonu, které chceme
studovat 1. Prednáška
Stažení homologních sekvencí od relevantních
taxonu z databáze 2. prednáška
Tvorba alignmentu 3. prednáška
Bude náplní prvního praktika 30. 11. 900
10
alignment
Mutliple sequence alignment (MSA)
Kontrolní otázka Co v alignmentu predstavuje
jeden znak?
11
FINGERPRINT
Kontrolní otázka Co ve fingerprintu predstavuje
jeden znak?
12
ODHAD GENETICKÉ VZDÁLENOSTI
13
Distance z podobnosti vzoru
Koeficient genetické vzdálenosti dle. Nei a Li
1979, PNAS 76, 1979 Pro každou dvojici (x, y)
spocteme všechny fragmenty (Mx, My) a dále
fragmenty vyskytující se v obou
elektroforetogramech ( Mxy) Vypocteme podíl
shodných fragmentu I 2Mxy/(Mx My) a z nej
distanci D 1- I
RAPD/RFLP
X Y
14
Distance z frekvence alel
Rogersova vzdálenost (pro všechny alely i lokusu
v populacích XA a XB) D (0,5 S(xAi - xBi)2)0,5
Frekvence alel v jednom lokusu
Alela Populace A Populace B
1 0,12 0,20
2 0,48 0,30
3 0,40 0,50
D (0,5((0,12-0,20)2(0,48-0,30)2(0,40-0,50)2))0,
5 (0,5(0,00640,03240,01))0,5 0,156
15
Distance z frekvence alel
Rogersova vzdálenost (pro všechny alely i lokusu
v populacích XA a XB) D (0,5 S(xAi - xBi)2)0,5
Vzdálenost Cavali-Svorza a Edwardse (1967) (pro
všechny alely u lokusu v populacích X a Y)
V prípade, že použijeme více lokusu bude celková
vzdálenost prumerem vzdáleností lokusu.
Reynoldsova distance (1983) nebo Neiova distance
(1972, 1978) berou v poraz biologické procesy.
16
VÝPOCET p
p podíl rozdílných nukleotidu mezi sekvencemi 2
taxonu
p nd/n
AATGTAGGAATCGC ACTGAAAGAATCGC
p 3/14 0,21
Odhad p z reasociacní kinetiky DNA-DNA
hybridizace
p ?Tm . 0,01 (0,015)
17
FREKVENCE SUBSTITUCÍ JE VYŠŠÍ NEŽ p
ACTGAACGTAACGC
C T
T C
G
T
C T
Koincidence
Zpetná substituce
Jednoduchá substituce
Vícenásobná substituce
K 12, p 3
Vidíme jen 3 rozdíly (p3/14), ale ve skutecnosti
došlo ke dvanácti substitucím (D 12/14).
18
SUBSTITUCNÍ SATURACE
0,95 PROTEINY
Pocet pozorovaných rozdílu
0,75 DNA
p
P distance není aditivní
Skutecný pocet substitucí na jednu pozici
19
ODHAD POCTU SUBSTITUCÍ
ut
Sekvence A
Sekvence B
Substitucní rychlost
Casový interval
Sekvence A - AATGTAGGAATCGC
Sekvence B - ACTGAAAGAATCGC
20
Jukes-Cantor príklad korekce na mnohonásobné
substituce
u substitucní rychlost u/3 rychlost
substituce za jednu konkrétní bázi
(napr. A -gt G)
Rychlost substituce za kteroukoli bázi
i sebe sama4/3 u
Ocekávaný pocet substitucí za cas t4/3 ut
u/3
21
Poissonovo rozdelení Rozdelení poctu výskytu málo
pravdepodobných jevu
25 let ? 20 substitucípro 5 let ?4
8
?4
?4
?4
Pravdepodobnost, že k žádné události nedojde
(k0) je e -?
Pravdepodobnost, že dojde práve ke k událostem je
f(k,?) (?k e-?)/k!
22
Jukes-Cantor príklad korekce na mnohonásobné
substituce
Pravdepodobnost, že za cas t k žádné události
nedojde e -4/3 ut
Pravdepodobnost, že za cas t k události dojde
1- e -4/3 ut
Pravdepodobnost, že dojde ke konkrétní událostiP
(CA) 1/4 (1- e -4/3 ut)
u/3
Pravdepodobnost, že dojde ke zmeneDs 3/4 (1- e
-4/3 ut)
Ocekávaný pocet substitucí za cas t4/3 ut
23
Jukes-Cantor príklad korekce na mnohonásobné
substituce
Pravdepodobnost, že dojde ke zmeneDs 3/4 (1- e
-4/3 ut)
ut
Sekvence A
Sekvence B
Substitucní rychlost
Casový interval
Sekvence A - AATGTAGGAATCGC
Sekvence B - ACTGAAAGAATCGC
p pocet neshodných míst/ délka sekvence je
odhadem Ds.
24
Jukes-Cantor príklad korekce na mnohonásobné
substituce
Pravdepodobnost, že dojde ke zmeneDs 3/4 (1- e
-4/3 ut)
Korigovaný pocet substitucíD ut -3/4 ln(1-
4/3 p)
Rozptyl D
V(D) (p(1 -p))/(L(1 - 4/3 p)2) L délka
sekvence
u/3
25
Jukes-Cantor príklad
Korigovaný pocet substitucíD ut -3/4 ln(1-
4/3 p)
Príklad naší sekvence D -3/4 ln(1- 4/3
0,21) D 0,246
u/3
26
Jukes-Cantor Obecne použitelné odvození
G
C
T
A
-u
u/3
u/3
u/3
A
u/3
G
u/3
u/3
-u
C
u/3
-u
u/3
u/3
-u
u/3
u/3
u/3
T
4/3 ut
4/3 ut
Pravdepodobnost, že dojde ke konkrétní událostiP
(CA) 1/4 (1- e -4/3 ut)
27
Jukes-Cantor Soucet rádku je 1
G
C
T
A
-u
u/3
u/3
u/3
A
u/3
G
u/3
u/3
-u
C
u/3
-u
u/3
u/3
-u
u/3
u/3
u/3
T
Suma 1
4/3 ut
4/3 ut
28
Jukes-Cantor Nekonecne vzdálené sekvence se
podobají z 1/4
ut8
Sekvence A
Sekvence B
4/3 ut
4/3 ut
29
Jukes-Cantor Je symetrický
G
C
T
A
-u
u/3
u/3
u/3
A
u/3
G
u/3
u/3
-u
C
u/3
-u
u/3
u/3
-u
u/3
u/3
u/3
T
Sekvence A
Sekvence B
30
Jukes-Cantor Je symetrický
G
C
T
A
-u
u/3
u/3
u/3
A
u/3
G
u/3
u/3
-u
C
u/3
-u
u/3
u/3
-u
u/3
u/3
u/3
T
Sekvence A
Sekvence B
31
DALŠÍ MODELY
a
Kimura 2P Kimura 2 parametrový
32
DALŠÍ MODELY
a
Kimura 2P Kimura 2 parametrový
33
DALŠÍ MODELY
Kimura 2P Kimura 2 parametrový
a
D 0,5 ln(a) 1/4 ln(b) a 1/(1 - 2P -
Q) b 1/(1 -2Q)
P podíl transic Q podíl transverzí
Rozptyl D
V(D) a2P c2Q -(aP cQ)2/L c (a
b)/2 L délka sekvence
34
DALŠÍ MODELY
Kimura 2P Kimura 2 parametrový
a
D 0,5 ln(a) 1/4 ln(b) a 1/(1 - 2P -
Q) b 1/(1 -2Q)
Príklad naší sekvence P2/140,14 Q1/140,07 a
1/(1 20,14 0,07) 1,54 b 1/(1 -20,07)
1,16 D 0,5 ln(1,54) 1/4ln(1,16)0,254
35
DALŠÍ MODELY
F84
Rovnovážné frekvence nukleotidu pA pC pG pT
36
DALŠÍ MODELY
GTR General time reversible
?
Rovnovážné frekvence nukleotidu pA pC pG pT
Parametry frekvence (rychlost) zámen (aß?de?) a
frekvence nukleotidu (pA pC pG pT) se odvozují z
analyzovaných sekvencí.
37
RUZNÉ MODELY FUNGUJÍ RUZNE
Odhad poctu substitucí
Skutecný pocet substitucí
38
LogDet distance
dxy -ln (det Fxy)
Sekvence A
Sekvence B
Alignment 900 pozic
Fxy
0,249 0,006 0,027 0,009 0,003 0,166 0,001 0,018 0,
027 0,006 0,256 0,004 0,006 0,021 0,009 0,194
dxy -ln (det Fxy) -ln (0.002) 6,216
39
JAKÉ DISTANCE POUŽÍVAT?
Modely s vetším poctem parametru (GTR) jsou
flexibilnejší a vetšinou presnejší než metody
jednoduché. Potrebují však velké množství
parametru a distance jimi vypocítané mají vetší
rozptyl. Pro kratší úseky proto dávají horší
výsledky. Simulace ukázala, že pro Dlt 0,5
dávají pomerne presné výsledky i nejjednoduší
metody (Jukes-Cantor, Kimura). Pro vetší
distance (a dostatecne dlouhé sekvence) je lépe
použít složitejší modely (GTR).
Recommended
CrystalGraphics Presentations





























