
Photosynthesis: light reactions and carbonassimilation reactions dark reactions - PowerPoint PPT Presentation
1 / 27
Title:
Photosynthesis: light reactions and carbonassimilation reactions dark reactions
Description:
Photosynthesis: light reactions and carbon-assimilation ... Stoichiometry of CO2 assimilation in the Calvin cycle. for every three CO2 molecules fixed: ... – PowerPoint PPT presentation
Number of Views:2827
Avg rating:5.0/5.0
Title: Photosynthesis: light reactions and carbonassimilation reactions dark reactions
1
Photosynthesis light reactions and
carbon-assimilation reactions (dark reactions)
171
2
In higher plants, photosynthesis takes place in
choloroplasts
- Similarity to mitochondria
- Two membranes
- Permeable outer membrane
- Selective inner membrane
- Difference to mitochondria
- Inner membrane forms a compartment that contains
flattened membranous sacs called thylakoids - (not contiguous with the inner membrane)
- Thylakoids are arranged in stacks called grana
- Thylakoid membranes contain
- photosynthetic pigments
- enzymes of photosynthesis
- Stroma
- aqueous phase enclosed by the inner membrane
- contains most of the enzymes of the carbon
assimilation reactions
172
3
Photosynthesis background
1937 Robert Hill determined that leaf extracts
containing chloroplasts could reduce a
nonbiological electron acceptor (Hill Reagent)
and make O2
2-6, dichlorophenolindophenol (a Hill reagent) is
blue when oxidized and colorless when
reduced The first evidence that absorbed light
energy causes electrons to flow from H20 to an
electron acceptor Hill also determined that C02
was neither required or reduced under conditions
that generated O2
1940 Severo Ochoa showed that NADP is the
biological electron acceptor in chloroplasts
173
4
- When a photon of light is absorbed
- an electron in the absorbing molecule is lifted
to a higher energy level - A molecule that has absorbed a photon is in an
excited state (unstable) - An electron lifted into a higher energy orbital
usually returns rapidly to its normal lower
energy orbital or ground state (stable) - As the electron returns to its ground state it
gives up the absorbed quantum - as light (fluorescence)
- as heat
- as chemical work
- In photosynthesis an alternative mode of decay
exists that involves direct transfer of
excitation energy from an excited molecule to a
neighboring molecule
174
5
Chlorophylls absorb light energy for
photosynthesis
- Green pigments with polycyclic, planar
structures resembling the protoporphyrin of
hemoglobin - Mg2 (not Fe2)
- All chlorophylls have a long phytol side chain
esterified to a carboxyl group in ring IV - The four inward oriented nitrogen atoms are
coordinated with the Mg2 - Polyene structure with alternating single and
double bonds characteristically show - strong absorption in the visible region of the
spectrum
175
6
Chlorophylls are not the only light absorbing
molecules in plants
176
7
Absorption of visible light by photopigments
Plants are green because their pigments absorb
light from the red and blue regions of the
spectrum (all higher plants contain chlorophyll a
and b) This leaves primarily green light to be
reflected or transmitted The relative amounts of
chlorophylls and accessory pigments are
characteristic for different plants and the
variation in the proportions of these pigments is
responsible for the range of colors of
photosynthetic organisms
177
8
Chlorophyll is always associated with specific
binding proteins forming Light Harvesting
Complexes (LHCs)
- Light Harvesting Complex II
- Seven molecules of chlorophyll a (in green)
- Five molecules of chlorophyll b (in red)
- Two molecules of Lutein (an accessory pigment)
- (in yellow)
- The functional unit is a trimer of 36 chlorophyll
- and six Lutein molecules
Accessory pigments extend the range of light
absorption
- In addition to chlorophylls, the thylakoid
membranes contain secondary light-absorbing
pigments (accessory pigments) called carotenoids - lutein (yellow)
- Carotene (red-orange)
178
9
Chlorophyll Funnels Absorbed Energy to Reaction
Centers by Exciton Transfer
Photosystem functional arrays of light absorbing
pigments of the thylakoid (or bacterial) membranes
All the pigment molecules in a photosystem can
absorb photons, but only a few chlorophyll
molecules associated with the photochemical
reaction center are specialized to transduce
light into chemical energy
The other pigment molecules (the ones that
absorb but dont transduce light) are called
light-harvesting or antenna
molecules They absorb light energy and transmit
it rapidly and efficiently to the reaction center
Note also embedded in the thylakoid membrane is
ATP synthase
179
10
Exciton Transfer
When isolated chlorophyll molecules in vitro are
excited by light the absorbed energy is quickly
released as fluorescence and heat But when
chlorophyll in intact leaves is excited by
visible light (step 1) there is very little
fluorescence Step 1 may be repeated a number of
times between successive antenna molecules until
a reaction center chlorophyll is reached
1710
11
There are two photosystems in plants
The combined activities of the two plant
photosystems move electrons from water to
NADP This way, some of the energy of absorbed
light is conserved as NADPH Simultaneously
protons are pumped across the thylakoid membrane
and energy is conserved as an electrochemical
potential and this proton gradient drives the
synthesis of ATP (the other energy conserving
product of the light reactions) photophosphorylat
ion
1711
12
Pathway of electron transfer from H2O to NADP
To raise the energy of electrons derived from H2O
to the energy level required to reduce NADP each
electron must by lifted twice by photons
absorbed in PSI and PSII One photon is required
per electron in each photosystem After
excitation, the high energy electron flows
downhill through the components of each
photosystem Protons move across the thylakoid
membrane during the water splitting reaction and
during electron transfer through the cytochrome
b6f complex This creates the proton gradient
required for ATP synthesis
1712
13
Photosystem II splitting water
A photon is absorbed by the light-harvesting
system of PSII The photon is funneled to a
reaction center chlorophyll (P680) Excitation of
P680 raises the molecule from the ground state to
an excited state (at -0.8V) Excited P680 is now
a good reducing agent and can transfer an
electron to a lower-energy electron acceptor
pheophytin a This electron is then transferred
through a series of molecules that make up PSII
to photosystem I When P680 (of PSII) gives up
an electron to pheophytin it must acquire an
electron to return to its ground state in
preparation for capture of another photon
Theoretically, this electron could come from any
number of sources (organic or inorganic) Photosynt
hetic bacteria use acetate, succinate, malate or
sulfide (depending on what is available in a
particular ecological niche) 3 billion years
ago primitive photosynthetic bacteria developed a
photosystem capable of taking electrons from a
donor that was always available water In this
process two water molecules are split yielding
four electrons, four protons and molecular
oxygen A single photon of visible light does not
have enough energy to break the bonds in
water four photons are required
1713
14
The four electrons abstracted from water do not
pass directly to P680
P680 can accept only one electron at a
time Oxygen Evolving Complex (water-splitting
complex) passes four electrons one at a time to
P680 This complex contains a cluster of four
oxygen bridged Manganese atoms (MnC) which can
exist in a series of oxidation states (from 2 to
7) so a cluster of 4 Mg ions can definitely
donate or accept four electrons (note this
complex has not yet been purified so mechanisms
are speculative) The sequential absorption of
four photons, each causing the loss of one
electron from the Mn center produces and
oxidizing agent that can take four electrons from
two molecules of H20 and produce 02 The
electrons lost from the Mn center pass one at a
time to an oxidized Tyr residue in P680 The four
protons are released into the thylakoid lumen
1714
15
Cytochrome b6f complex links photosystems II and I
electrons produced as a result of the excitation
of P680 in PSII are carried to P700 of PSI via
the cytochrome b6f complex (cytochrome f complex)
Like complex III in mitochondria cytochrome b6f
complex conveys electrons from a reduced quinone
(PQB in chloroplasts) to a water soluble protein
that carries one electron (plastocyanin in
chloroplasts) In the Q cycle electrons pass one
at a time from PQBH2 to cytochrome b6 This cycle
results in the pumping of protons from the
stromal compartment to the thylakoid lumen (up to
four protons move for each pair of electrons
the exact number is not yet clear) The result is
the formation of a proton gradient across the
thylakoid membrane as electrons are passed from
PSII to PSI
1715
16
Photosystem I production of NADPH
For every two photons absorbed (one by each
photosystem) one electron is transferred from H20
to NADP To form one molecule of O2 (which
requires the transfer of four electrons from two
H20 to NADP ) a total of eight photons must be
absorbedfour by each photosystem
In PSI, the excited reaction center P700 loses an
electron to an acceptor (A0) This results in a
charge separation at the photochemical reaction
center P700 is a strong oxidizing agent which
aquires an electron from plastocyanin (soluble
Cu-containing electron transfer protein A0- is a
very strong reducing agent that passes its
electron through a series of electron carriers
that lead to NADP, reducing it to NADPH Protons
are pumped from the stroma into the thylakoid
lumen but exactly how many is uncertain because
its not clear how many are pumped by the b6f
complex estimated 12 protons pumped per O2
produced
1716
17
ATP synthesis by photophosphorylation
1717
18
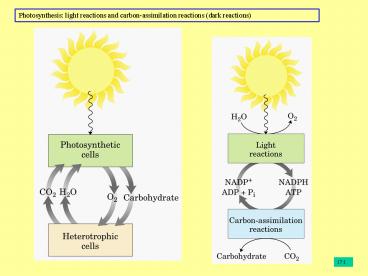
Photosynthetic Carbohydrate Synthesis
The precursor molecules for synthesis of
carbohydrates in animal cells always have at
least three carbons all of which are less
oxidized than the carbon in C02
Photosynthetic organisms can synthesize
carbohydrate from C02 and water, reducing C02
using energy from the ATP and NADPH generated by
photosynthetic electron transfer
autotrophic organisms (phototrophic or
chemotrophic) can use CO2 as the sole source of
all the carbon atoms required for the
biosynthesis of cellulose and starch, lipids and
proteins heterotrophic organisms can incorporate
CO2 into organic compounds in some
reactions (pyruvate carboxylase reaction of
gluconeogenesis) (acetyl-CoA carboxylase reaction
of fatty acid synthesis) (carbamoyl phosphate
synthetase I reaction of urea formation) but the
CO2 is lost in a subsequent reaction step
1718
19
Carbon assimilation reactions carbon fixation
(dark reactions)
1719
20
Step1 Fixation of CO2 into 3-phosphoglycerate
The substrates are ribulose 1,5-bisphophate
CO2 the enzyme is ribulose 1,5-bisphosphate
carboxylase/ oxygenase RUBISCO
The most abundant enzyme in the biosphere
eight large subunits (each contains an active
site) eight small subunits (function unknown)
one of the 3-phosphoglycerate molecules contains
the carbon from CO2
1720
21
Step 2 converstion of 3-phosphoglycerate to
glyceraldehyde 3-phosphate
This two step reaction is essentially a reversal
of the corresponding reactions in
glycolysis (except that NADPH is the cofactor
instead of NADH) the glyceraldehyde 3-phosphate
is either converted to startch in the cholorplast
and stored for later use or transported to the
cytosol and converted to sucrose for transport to
growing regions of the plant
In developing leaves, a significant portion may
also be degraded by glycolysis to provide
additional energy for growth
1721
22
Step 2
1722
23
Step 3 Regeneration of ribulose 1,5
bisphosphate from triose phosphates
1723
24
- for every three CO2 molecules fixed
- one triose phosphate is produced
- nine ATP are consumed
- six NADPH are consumed
Stoichiometry of CO2 assimilation in the Calvin
cycle
The source of the ATP and the NADPH is the
light-driven reactions of photosynthesis
In the dark, the production of ATP and NADPH
ceases as does the incorporation of CO2
All the reactions of the Calvin cycle except
those catalyzed by RUBISCO, sedoheptulose and
ribulose 5-phosphate kinase also take place in
animal tissue
1724
movie 245601
25
RUBISCO ribulose 1, 5 bisphosphate
carboxylase/oxygenase
1725
Apparently when RUBISCO evolved, oxygen was not a
significant component of the atmosphere and the
active site of RUBISCO does not discriminate well
between CO2 and O2 Therefore, RUBISCO also
functions as an oxygenase
The condensation of O2 with ribulose 1, 5
bisphosphate forms phosphoglycolate a
metabolically useless product Salvaging the
phosphoglycolate( the glycolate pathway) requires
the input of energy This process is called
photorespiration ( O2 is consumed and CO2 is
produced)
The oxygenase activity of RUBISCO occurs
concurrently with the carboxylase activity and
can constitute a significant waste of energy
In addition the affinity of rubisco for CO2
decreases when temperature increases (making
tropical plants even more susceptible to
photorespiration
26
Some plants have a mechanism to minimize
photorespiration
The Hatch-Slack Pathway (C4 cycle) occurs
primarily in plants that grow in the
tropics (because the affinity of RUBISCO for CO2
is decreased by heat)
1726
27
1727
Recommended
CrystalGraphics Presentations





























